
|
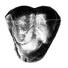
Neolicaphrium recens Frenguelli,1921,the only surviving proterotheriidae (Litopterna, Mammalia) into the south american Pleistocene.
Published online: 30/07/2001
Keywords:
Litopterna; Neolicaphrium recens; Pleistocene; Proterotheriidae; South America
Abstract
The litoptem Proterotheriidae are extinct endemic South American ungulates frequently used as an example of evolutionary convergence with the horses. They were considered to be exclusively Tertiary representatives with the youngest record being in the late Pliocene, before the appearence of the equids and cervids during the Great American Interchange. Two undoubted Pleistocene records in Argentina and the specimen here described from Uruguay, confirm the persistence of the proterotherids into that period. In the Quaternary, these ungulates are found outside the typical pampean region and probably were confined to a few northern and warmer more forested relictual microhabitats.
PV article infos
Published in Vol. 30, Fasc. 1-2 (2001)
|
PDF
|
|
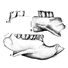
Carolocoutoia ferigoloi nov. and sp. (Protodidelphidae), a new Paleocene "opossum-like" marsupial from Brazil.
Published online: 15/12/1998
Keywords:
Brazil; Didelphimorphia; Itaborai; Marsupialia; New taxa; Paleocene; Protodidelphidae; South America
Abstract
Carolocoutoia ferigoloi gen. et sp. nov. is the largest of protodidelphid marsupials, known from Middle Paleocene levels at Itaboraí Formation, southeastern Brazil. It differs from other members of this family in having molars with low cusps which are basally inflated, rounded crests without cutting edges, and a thick enamel layer which is wrinkled, specially at the labial half. A comparative analysis among representatives of this family led us to recognize only three genera undoubtely assignable to it: Protodidelphis PAULA COUTO, 1952, Robertbutleria MARSHALL, 1987, and Carolocoutoia gen. nov. Protodidelphids lack the basic derived features diagnostic of Polydolopimorphian marsupials, while most of its derived features agree with its belonging to the Didelphimorphia. Protodidelphids comprise a specialized clade of opossum-like marsupials adapted to frugivorous or frugivore-omnivorous feeding habits. They differ from other didelphimorphians in having very large, spire-like entoconids, reduced and antero-posteriorly compressed paraconids, absence of stylar cusp C and of para- and metaconules, large stylar cusps B and D which are proximate to each other, short postmetacristae, eccentric protocones, and molars that increase rapidly in size from M/ml to M/m3.
PV article infos
Published in Vol. 27, Fasc. 3-4 (1998)
|
PDF
|
|
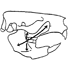
Octodontid-like Echimyidae (Rodentia) : an upper Miocene episode in the radiation of the family
Published online: 20/05/1994
Keywords:
Argentina; Echimyidae; Miocene; New taxa; Rodentia; South America
https://doi.org/10.18563/pv.23.1-4.199-210
Abstract
Reigechimys octodontiformis gen. et sp. novo and R. plesiodon sp. novo are described. They represent the frrst record of the family Echimyidae for the Cerro Azul Formation (Huayquerian Age, Late Upper Miocene) at La Pampa Province, central Argentina. Both species have hypsodont cheek teeth with an eight-shaped occlusal design. This dental morphology represents a noticeable case of convergence to octodontids and indicates that these echimyids inhabited open environments.
PV article infos
Published in Vol. 23, Fasc. 1-4 (1994)
|
PDF
|
|
Schmelzmikrostruktur in den inzisiven alt-und neuweltlicher histricognather nagetiere
Published online: 15/12/1992
Keywords:
Africa; Caviomorpha; Ctenodactyloidea; Deseadan; Enamel microstructure; Hunter-Schreger bands; Hystricognathi; Incisors; Ischyromyoidea; multiserial; Paleobiogeography; pauciserial; Phiomorpha; Rodentia; South America
https://doi.org/10.18563/pv.21.ext.1-168
Abstract
Enamel microstructure in the incisors of Old- and New World hystricognath rodents:
The incisor enamel microstructure in more than 100 genera of fossil and Recent hystricognath and sciurognath rodents was studied. A multiserial schmelzmuster is present in the Hystricognathi, the Ctenodactylidae, advanced Chapattimyidae, and in Pedetes. A redefinition of pauciserial and multiserial HSB is given that makes the two enamel types unambiguously distinguishable which apparently represent well defined evolutionary levels. In the pauciserial Schmelzmuster the IPM is thicker than in the multiserial one. In pauciserial HSB the IPM always surrounds each prism, and the crystallites of the IPM run parallel to prism direction; transition zones between HSB are lacking; the inclination of the HSB is normally very low and the prism cross sections are not flattened but somewhat irregular. The number of prisms per HSB is no good distinctive character for pauciserial and multiserial HSB, since there exists a wide overlap. The pauciserial schmelzmuster is primitive, the multiseiial derived because: 1. the pauciseiial schmelzmuster appears earlier in the fossil record in the most primitive rodents (Paramyids s.l. and Ctenodactyloids); 2. the Eocene Ctenodactyloidea show pauciserial HSB but the Oligocene and younger ones are characterized by multiserial HSB; 3. in the outgroup comparison, the Eurymylidae (Mixodontia) show pauciserial HSB; 4. biomechanically, multiserial HSB strenghten the enamel better than pauciserial HSB, since their IPM runs nearly always in an angle of 45° or more to the prisms.
In multiseríal HSB three subtypes can be distinguished which are differentiated by the IPM orientation. Primitive is a (rarely strict) parallel or acute angular, anastomozing IPM, and derived is an interrow sheet-like ("plattenartige") IPM. This evolutionary polarity is indicated by enamel evolution in the Ctenodactylidae which show an acute angular IPM in the Oligocene and a rectangular interrow sheet-like IPM since the Miocene. Among the Caviomorpha a rectangular interrow sheet-like IPM is restricted to the Octodontoidea; therefore they must be considered derived in terms of their enamel structure. The first multiserial HSB in rodent incisors appear in phiomyids or chapatrimyids from the Upper Eocene of Algeria. The IPM is acute angular and anastomozing. The worldwide next younger multiserial HSB are found in Lower Oligocene phiomyids of Fayum, Egypt There already a rectangular interrow sheet like IPM is present (in Metaphiomys) besides the acute angular anastomozing IPM.
The first Caviomorpha from the Deseadan (Oligocene-Miocene) likewise show already acute angular anastomozing IPM (e.g. Scozamys) and rectangular interrow sheet-like IPM (Platypittamys). Therefore the first Caviomorpha cannot be positioned close to a transition from pauciserial to multiserial HSB. In none of the potential caviomorph ancestors from southern North America multiserial HSB or transitional stage between pauciserial and multiserial HSB could be found. The similarities between the enamel types of the Fayum rodents and the rodents from the Deseadan of South America make a derivation of the Caviomorpha from Paleogene North African phiomorph rodents or their direct ancestors most probable. This supports at the same time a descent of the platyrrhine Primates from North African anthropoids.
PV article infos
Published in Vol. 21, Ext (1992)
|
PDF
|

|
Analysis of changing diversity patterns in Cenozoic land mammal age faunas, South America
Published online: 30/03/1990
Keywords:
Cenozoic; Chronofaunas; diversity; Equilibrium theory; Extinction; Land mammal faunas; Origination; South America
https://doi.org/10.18563/pv.19.4.169-210
Abstract
Comparison of various measurements of taxonomic evolution using stratigraphic range data for orders, families and genera of land mammals indicates several means by which deficiencies of the South American fossil record (e.g., presence of hiatuses, unequal temporal and geographic representation of ages, unequal systematic treatment) may be normalized, thus permitting a less distorted appreciation of diversity pattern and trend. Initial radiation of native taxa resulted in a relative equilibrium by early Eocene time. Subsequent increases in absolute diversity were apparently induced by immigration at the family level and by environmental factors at the generic level. Miocene through Pleistocene phases of faunal stability, herein characterized as chronofaunas, are punctuated by rapid turnover events resulting from a complex of factors, including adaptive radiation of immigrant taxa into unoccupied eco-space; environmental and concomitant habitat change induced by orogenic events of the Andes; and biotic interactions between native and immigrant taxa, including competition and prey naivete. The first two factors account for major faunal transitions in the South American middle and late Tertiary; immigration-induced turnover may have been of greater importance in shaping the character of the fauna upon the Great American Interchange and the arrival of man in the Neotropics
PV article infos
Published in Vol. 19, Fasc. 4 (1990)
|
PDF
|
|
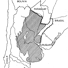
Mammals and stratigraphy of the continental mammal-bearing Quarternary of South America
Published online: 16/12/1984
Keywords:
Geochronology; Mammalia; Quaternary; South America; Stratigraphy
https://doi.org/10.18563/pv.14.ext
Abstract
Previous chronological arrangements of South American Quaternary land mammal faunas are appraised on the basis of current geological and paleontological data. Three South American late Pliocene-Pleistocene land mammal ages are conventionally recognized, from oldest to youngest, the Uquian, Ensenadan, and Lujanian ; all are defined on Argentine faunas.
The Uquian is based fundamentally and historically on the fauna from the Uquía Formation in Jujuy Province, northwestern Argentina. Important known formations in Argentina yielding Uquian Age faunas include the sub-surface Puelche Formation (or Puelchense) near the city of Buenos Aires, and the Barranca de Los Lobos and Vorohué Formations between Mar del Plata and Miramar, Buenos Aires Province. A tentative subdivision is propos-ed for the Uquian into three subages based on knowledge of the Mar del Plata-Miramar sequence, from oldest to youngest, the Barrancalobian, Vorohuean, and Sanandresian. In Argentina the Uquian is presently marked by the first known record of Scelidodon, Hydrochoeropsis, Ctenomys, Canidae, Ursidae, Gomphotheriidae, Equidae, Tapiridae, Camelidae, Cervidae, and the last known record of Thylatheridium, Thylophorops, Dankomys, Eumysops, Pithanotomys, Eucoelophorus, Hegetotheriidae, Sparassocynidae, and Microtragulidae.
The Ensenadan Age is based on the fauna from the Ensenada Formation near the city of Ensenada, Buenos Aires Province. In Argentina the Ensenadan is marked by the first known record of Lomaphorus, Neothoracophorus, Plaxhaplous, Cavia, Lyncodon, Lutra, Galera, Smilodon, Dicotyles, Lama, Vicugna, the last known record of Orthomyctera, and the only known record of Brachynasua.
Typícal beds of late Lujanian Age in Argentina consist of fluvial deposits occupying stream channels, and shallow basins, often incised into beds of early Lujanian (i.e. Bonaerian of early workers) and Ensenadan Age. The Lujanian Age is based on a fauna from beds along the Rio Luján, about 65 km west of the city of Buenos Aires, Buenos Aires Province. The Lujanian in Argentina is marked by the first record of Equus, Chlamyphorus, and Holochilus, and the last record of Megatherioidea, Glyptodontoidea, Arctodus (=Arctotherium), Smilodon, Litopterna, Notoungulata, Proboscidea, Equidae, Morenelaphus, and Palaeolama.
These land mammal ages are often difficult to recognize in other South American countries. The compositions of South American Pleistocene faunas vary with the environment. Some taxa were widely distributed in fossil deposits throughout the continent, but their occurrences need not reflect synchroneity. This is a result of changing climates and habitats in time. Consequently, proposed intracontinental correlations need confirmation based on magnetostratigraphy and a radioisotope time scale. Paleontologic characterizations of these land mammal ages (i.e. first and last record, and guide fossils) are useful for much of Argentina, but extensions to most of the other parts of South America are at best tenuous.
The majority of known non-Argentine Pleistocene faunas are believed to be Lujanian in age. Possible non Argentine early Pleistocene (Uquian) faunas include Ayo Ayo and Anzaldo in Bolivia, and Cocha Verde in southern Columbia. A possible middle Pleistocene (Ensenadan or early Lujanian) fauna is the Chichense of Ecuador. Paleomagnetic and radioisotopic date (MacFadden et al., 1983) clearly indicate that the greater part of the Tarija fauna (Bolivia) is Ensenadan in age.
The end of the Pleistocene and beginning of the Holocene in South America is marked by extinction of nearly all large mammalian herbivores and their specialized large predators. Radiocarbon age determinations suggest that large scale extinctions of megafauna occurred between 15,000 and 8,000 yrs. B.P. (years before present).
PV article infos
Published in Vol. 14, Ext (1984)
|
PDF
|
|
Mammals and stratigraphy : Geochronology of the continental mammal-bearing Tertiary of south America.
Published online: 15/12/1983
Keywords:
Cenozoic; Geochronology; Mammalia; South America; Stratigraphy; Tertiary
https://doi.org/10.18563/pv.13.ext
Abstract
The principles and practices employed in establishment and recognition of South American land mammal ages are reviewed along with previous and present concepts of distinguishing time, rock, and faunal units. Previous chronological arrangements of South American Tertiary land mammal faunas are appraised on the basis of recent geological and paleontological data. Twelve South American Tertiary land mammal ages are here recognized [from oldest to youngest, Riochican (middle to late Paleocene); Casamayoran (early Eocene); Mustersan (middle Eocene); Divisaderan (late Eocene); Deseadan (early [to middle?] Oligocene); Colhuehuapian (late Oligocene); Santacrucian (early Miocene); Friasan (middle Miocene); Chasicoan (late Miocene); Huayquerian (latest Miocene); Montehermosan (early to middle Pliocene); and Chapadmalalan (late Pliocene)]. As all except the Friasian were originally defined on the basis of Argentine faunas, these are discussed first and at length, and each is reviewed with discussion of type locality, stratigraphy, type fauna, and faunal correlations. Non-Argentine faunas are then discussed country by country in alphabetical order.
A review is given of radioisotope dates obtained on volcanic rocks (i.e., basalts, tuffs) associated with mammalbearing beds in Argentina. Based on these age determinations and on correlation of the late Tertiary land mammals involved in the interchange between North and South America, a chronology of South American land mammal ages correlated with North American land mammal ages and European marine stages is proposed.
It is concluded that South America was an island continent through most of the Tertiary Period (ca 65 to about 3 Ma). As a result, the land mammal fauna of South America developed in isolation and was dominated by autochthonous endemic groups. Toward the end of the Tertiary (i.e., middle Miocene) a unique faunal balance had been achieved by the descendants of the ancient inhabitants (notoungulates, litopterns, condylarths, astrapotheres, edentates, marsupials) and of later (late Eocene) waif immigrants (caviomorph rodents, platyrrhine primates). A prominent feature of this mammal fauna was the combination of carnivorous and omnivorous marsupials with native placental herbivorous ungulates, subungulates, and edentates.
Sometime during the late Miocene, a limited but important interchange of mammalian taxa between North and South America took place. Procyonids (raccoons and their allies), a group of North American origin, first appear in South America in strata of Huayquerian Age, while members of the extinct South American ground sloth families Megalonychidae and Mylodontidae first appear in North America in early Hemphillian time. These groups dispersed along island arcs before the appearance of the Panamanian land bridge in the Pliocene (ca 3.0 Ma). Cricetine rodents, a group of North American origin, are first known in South America in strata of Montehermosan Age. The known taxa are too advanced and diversified to be considered the first of this group to invade South America. lt is believed by some workers that these rodents arrived before the Montehermosan, possibly in the late Miocene or earlier, by waif dispersal from North America.
The isolation of South America ended with the appearance of the Panamanian land bridge, which provided a direct, dry land connection between the two Americas. Across this portal an extensive interchange of terrestrial faunas occurred, and the fossil record documents an intermingling of these long-separated land mammals faunas.
The beginning of this interchange by land route in South America is marked by the appearance of mammals which evolved from North American emigrants in the Chapadmalal Formation of Argentina. These include a mustelid (Conepatus), a tayassuid (Argyrohyus), and four genera (Akodon, Dankomys, Graomys, Reithrodon) of cricetine rodents. The appearance of this contingent of northern animals favors the existence of the Panamanian land bridge by this time. Likewise, a large number of terrestrial vertebrates of South American origin appear in North America in beds of late Blancan Age date around 2.7 Ma. Among the mammals are Neochoerus, Erethizon, Glyptotherium, Glossotherium, Kraglievichia, and Dasypus.
PV article infos
Published in Vol. 13, Ext (1983)
|
PDF
|
|
Description des restes d'Elasmobranches (Pisces) du Dévonien moyen de Bolivie
Published online: 01/02/1977
Keywords:
South America
https://doi.org/10.18563/pv.7.4.126-132
Abstract
Some fragmentary remains of spines and endoskeletal elements, referred here to as ctenacanthid like elasmobranchs, are recorded in the Middle Devonian of Bolivia. These specimens, and some others from the Eodevonian of Brasil represent the only Devonian fish remains hitherto known from South America and indicate that further discoveries may be made in their original localities respectively.
PV article infos
Published in Vol. 07, Fasc. 4 (1977)
|
PDF
|

|
Rongeurs caviomorphes de l'Oligocène de Bolivie. 1 Introduction au deseadien de Bolivie
Published online: 01/08/1976
Keywords:
Rodentia; South America
https://doi.org/10.18563/pv.7.3.1-16
Abstract
The Tertiary of the Salla-Luribay basin consists of red beds affected by the second period of the andine compression, of Miocene ending age. The Tertiary layers are exposed at an approximate elevation of 3.500 to 4.000 meters. Two stratigraphic units can be distinguished in them: the Luribay conglomerates, in which vertical clifts result from erosion, and the Salla layers consisting mostly of consolidated clays. These clays are very fossiliferous and have furnished a rich vertebrate fauna which gave to R. Hoffstetter the possibility to establish the Oligocene age of these beds. Sediments of same age has been reported to be present in several other places of Bolivia, particularly near Lacayani, where have been collected highly hypsodont Rodents, different from those found in Salla-Luribay basin.
PV article infos
Published in Vol. 07, Fasc. 3 (1976)
|
PDF
|
|
Les vertébrés fossiles de Colombie et les problèmes posés par l'isolement du Continent sud-Américain.
Published online: 20/01/1969
Keywords:
Columbia; Cretaceous; Fauna; Quaternary; South America
https://doi.org/10.18563/pv.2.2.77-94
Abstract
A general view is given of the vertebrate faunas, Cretaceous to Quaternary of age, found in Columbia and of their principal characteristics. This view leads to the discussion of the isolation of the South American continent and of the role played by the Bolivar syncline with respect to North American immigrants during the Oligocene. The absence of marine deposits of Oligocene age in the north and northwest of Columbia suggests the possibility of a communication with Central America. This communication would have permitted the passage of hystricomorph rodents, of platyrrhine monkeys, and of colubrids. The non-occupation, until then, of the ecologie niches of these groups would have favored their installation beside the indigenous fauna. In this hypothesis it would no longer be necessary to admit that these vertebrates arrived as «island hoppers ››. The eco-biologic conditions would explain the absence of large-sized forms of North American origin.
PV article infos
Published in Vol. 02, Fasc. 2 (1969)
|
PDF
|
